Hầu hết các loại loạn nhịp tim là do bất thường của hình thành xung hoặc rối loạn dẫn truyền xung hoặc kết hợp cả hai. Ví dụ, Ngoại tâm thu thất khởi phát nhịp nhanh thất đơn dạng ở bệnh nhân nhồi máu cơ tim trước đó là do bất thường hình thành xung trong khi đó nhịp nhanh thất vào lại do sẹo là biểu hiện của bất thường dẫn truyền xung.
Hoạt động khởi kích hay lẫy cò và tự động tình bất thường được phân loại như rối loạn dẫn truyền xung.
HOẠT ĐỘNG KHỞI KÍCH
Hoạt động khởi kích là do sự phát xung của một nhóm tế bào cơ tim được khởi kích bỡi hàng loạt các xung trước. Hoạt động này được tạo ra bỡi một loạt hậu khử cực, là kết quả giảm điện thế màng. Điện thế màng dao động, khi đạt đến ngưỡng có thể hoạt hoá để tạo ra rối loạn nhịp đặc biệt này. Hậu khử cực mà xảy ra trước khi hoàn tất tái cực ( trong pha 2 hoặc pha 3 của điện thế hoạt động) được gọi là hậu khử cực sớm (EADs).Trong khi đó hậu khử cực mà xảy ra trong giai đoạn tái cực được gọi là hậu khử cực trì hoãn (DADs).
EAD được cho là nguyên nhân của rối loạn nhịp kèm với hội chứng QT dài bẩm sinh hay mắc phải. Nhịp tim chậm và khoảng ghép dài tạo điều kiện cho EAD xảy ra, ngược lại nhịp tim nhanh và khoảng ghép ngắn ức chế EAD. Hiệu quả magnesium ức chế phát triển EAD đã giải thích cho tác dụng của nó trong điều trị nhịp nhanh thất đa dạng thứ phát do kéo dài QT(xoắn đỉnh).
DADs là kết quả của dòng ion đi vào tạm thời mà khởi phát khử cực màng. Các dòng tạm thời này xảy ra để đáp ứng với tình trạng quá tải canxi nội bào và là hậu quả của sự phóng thích canxi từ mạng cơ tương. DAD củng do dòng vào được tạo ra bỡi sự trao đổi Na va Ca. DAD củng xảy ra ở các mô ngấm digoxin, và nhiều loạn loạn nhịp xảy ra kèm với digoxin được nghĩ là do cơ chế khởi kích hay lẫy cò. Nhịp tự thất tăng tốc sau nhồi máu cơ tim củng do quá tải canxi và DAD dẫn đến hoạt động khởi kích.
Biểu hiện lâm sàng: rối loạn nhịp mà do hoạt động khởi kích là loạn loạn nhịp xảy ra sau gia tăng nhịp xoang. Ví dụ thường gặp nhất là loạn nhịp thất từ buồng tống thất phải xảy ra trong bối cảnh gắng sức hoặc đáp ướng với các thuốc kích thích beta.
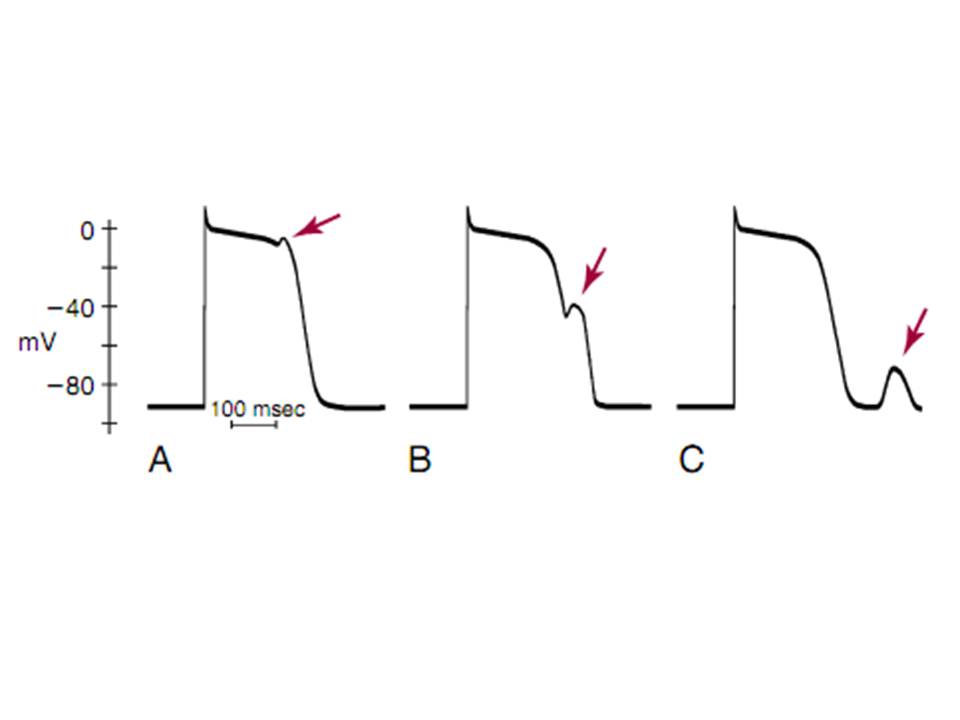
Hình 1. Các loại hậu khử cực. Hậu khử cực được chỉ bỡi các mũi tên. Điện thế hoạt động của tế bào purkinje với hậu khử cực sớm xảy ra ở pha 2(EADs) (A) và pha 3(EADs) (B), cũng như hậu khử cực trì hoãn (DADs) (C), xảy ra sau khi tái cực hoàn toàn.
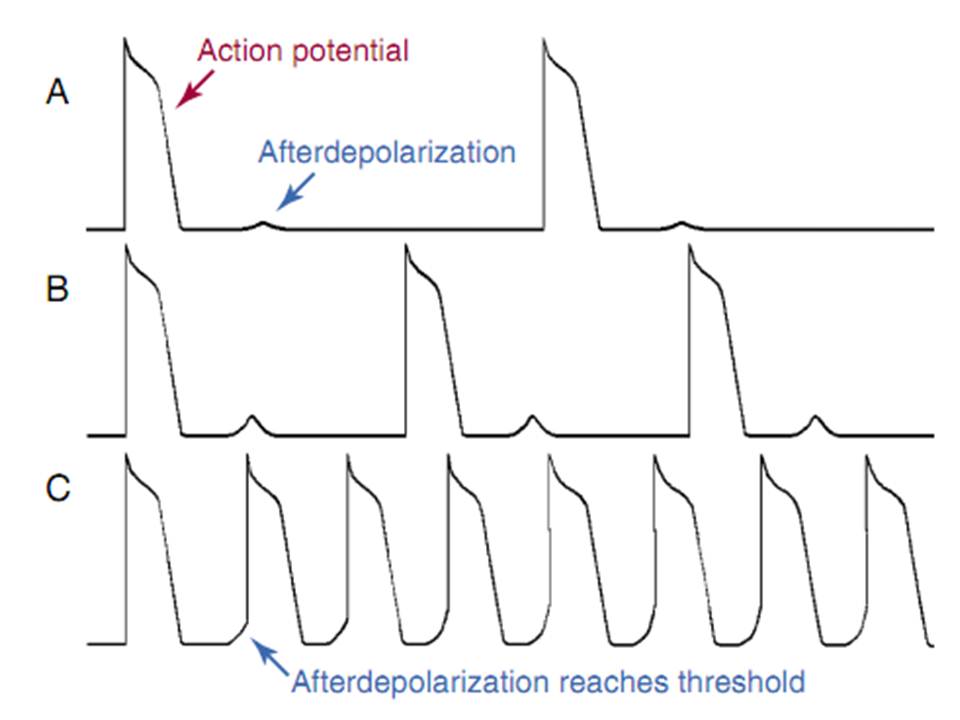
Hình 2. Hậu khử cực trì hoãn. A, hậu khử cực được thấy sau điện thế hoạt động ở nhịp chậm. B, khi nhịp nhanh hơn, hậu khử cực xảy ra sơm hơn và tăng biên độ. C, khi nhịp nhanh hơn, hậu khử cực xảy ra sớm hơn và đạt điện thế ngưỡng, dẫn đến phóng thích kéo dài.
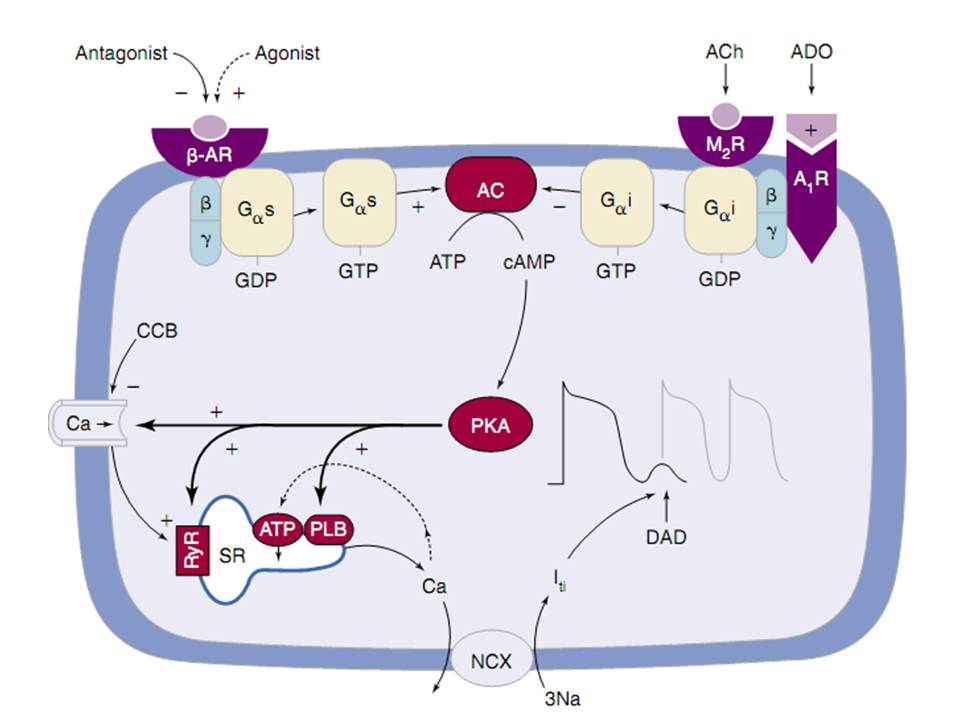
Hình 3. Biểu đồ dẫn truyền tín hiệu cho khởi đầu và chấm dứt hoạt động khởi kích qua trung gian AMP vòng.
TỰ ĐỘNG TÍNH BẤT THƯỜNG
Tim bình thường, tự động tính chỉ có ở nút xoang và mô dẫn truyền đặc biệt.Tế bào cơ nhĩ và thất hoạt động bình thường không co khử cực tâm trương tự phát và không có khởi phát xung tự phát, thậm chí khi các tế bào này không bị kích thích trong một khoảng thời gian dài do không có xung xâm nhập. Mặc dù các tế bào này củng có dòng ion tạo nhịp nhưng khoảng tạo nhịp của các dòng này ở các tế bào cơ nhĩ và thất thì âm hơn nhiều – 120 đến -170 mv so với sợi purkinje hay nút xoang. Do dó, trong khi điện thế màng lúc nghỉ sinh lý (-85 đến -95 mv) dòng ion tạo nhịp vẫn không hoạt động nên tế bào cơ thất không khử cực tự phát. Khi điện thế màng khi nghỉ của các tế bào này khử cực đủ đến khoảng -70 đến -30 mv lúc đó khử cực tâm trương tự phát có thể xảy ra và khởi phát xung lặp lại, một hiện tượng được gọi là tự động tính bất thường. Tương tự, các tế bào trong hệ thống purkinje có tự động tính bình thường ở mức điện thế màng cao sẻ có tự động tính bất thường khi điện thế màng giảm đền -60 mv hay thấp hơn, tự động tính bất thường này xảy ra ở vùng cơ tim thiếu máu. Khi điện thế màng của các sợi purkinje trong tình trạng ổn định giảm xuống khoảng -60 mv hay thấp hơn khi đó kênh If tham gia trong hoạt tính tạo nhịp bình thường ở các sợi purkinje bị đóng lại mất chức năng và vì vậy tự động tính không được gây ra bởi cơ chế tạo nhịp bình thường. Tuy nhiên, nó có thể được gây ra do cơ chế bất thường. Ngược lại, khi tự động tính của nút xoang, các chủ nhịp theo trình tự dưới nút xoang như nhĩ, nút nhĩ thất gia tăng được gây ra do cơ chế khác chứ không phải là sự gia tăng tự động tính bình thường, điều này chưa được chứng minh trên lâm sàng.
Điện thế màng ở mức thấp không phải là tiêu chuẩn duy nhất để xác định tự động tính bất thường.Vì giả tỉ nếu điều này đúng thì tự động tính của nút xoang củng được xem là bất thường. Vì vậy, một phân biệt quan trọng giữa tự động tính bình thường và bất thường đó là điện thế màng của các sợi thể hiện kiểu hoạt động bất thường được giảm từ mức bình thường riêng của nó. Vì lý do này, ví dụ khi tự động tính của nút nhĩ thất khi có điện thế màng ở mức thấp bình thường không được phân loại như là tự động tính bất thường.
Vài cơ chế khác nhau có thể gây ra tự động tính bất thường khi điện thế màng ở mức thấp bao gồm sự hoạt hoá và bất hoạt của dòng ion K tinh lọc trì hoãn, sự phóng thích canxi từ mạng cơ tương gây ra sự hoạt hoá cho dòng Na và dòng Ca đi vào(qua sự trao đổi của ion Na và Ca) và sự phân bố điện thế của dóng If. Người ta chưa chứng minh được loại nào trong số các cơ chế này được vận hành trong các tình trạng bệnh học khác nhau mà khi đó tự động tính bất thường có thể xảy ra.
Độ dốc pha 0 của điện thế hoạt động xảy ra tự phát do tự động tính bất thường có thể là do dòng vào của ion Na hoặc ion Ca hoặc kết hợp cả hai. Khi điện thế tâm trương khoảng -70 đến -50 mv, hoạt động lặp lại phụ thuộc vào nồng ion Na ngoại bào và hoạt động này có thể giảm hay xoá đi nhờ ức chế kênh ion Na. Khi điện thế tâm trương khoảng từ -50 đến – 30 mv kênh ion Na bị bất hoạt hoạt động lặp lại phụ thuộc vào nồng độ ion Ca ngoại bào và được giảm bỡi ức chế kênh ion Ca kiểu L.
Nhịp nội tại của ổ với tự động tính bất thường được quyết định qua điện thế màng, điện thế màng càng dương tự động tính càng nhanh. Vì vậy tự động tình bất thường ít nguy hiểm đến độ phải ức chế bằng tạo nhịp vượt tần số. Do đó, thậm chí khi nhịp xoang thấp cho phép các ổ ngoại tâm thu với tự động tính bất thường phát ra mà không cần khoảng dừng dài bắt buột trước đó.
Giảm điện thế màng tế bào cơ tim cần thiết cho tự động tình bất thường xảy ra có thể được tạo ra bỡi nhiều yếu tố liên quan bệnh tim, chẳng hạn như thiếu máu và nhồi máu cơ tim.Ví dụ, gia tăng nồng độ ion K ngoại bào xảy ra khi thiếu máu cơ tim có thể làm giảm điện thế màng; tuy nhiên tự động tính bình thường ở tế bào cơ nhĩ và thất sợi purkinje thường không xảy ra vì gia tăng dẫn truyền ion K do gia tăng nồng độ ion K ngoại bào. Catecholamin củng làm gia tăng tốc độ phát xung gây ra bỡi tự độnt ính bất thường vì vậy có thể góp phần vào sự dịch chuyển vị trí tạo nhịp, từ nút xong đến vùng tự động tính bất thường.
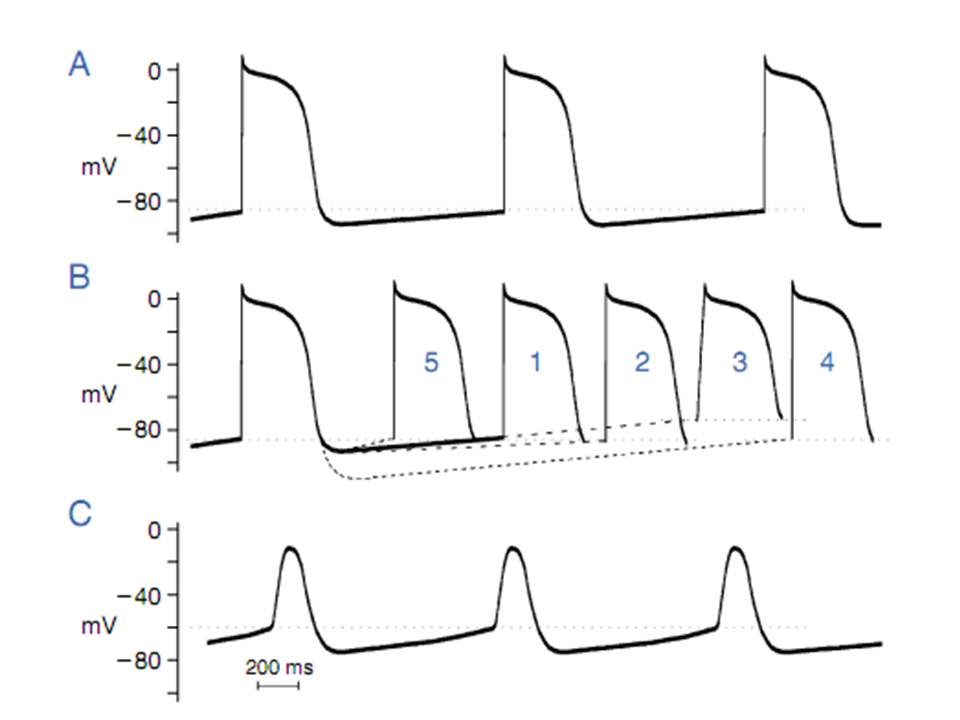
Hình 4. Bất thường tự động tính. A, điện thế hoạt động hệ thống his- purkinje bình thường.B, sự thay đổi tốc độ khử cực so vói cơ bản (1) chậm tốc độ khử cực pha 4, (2) tăng điện thế ngưỡng (3), bắt đầu từ điện thế màng khi nghỉ âm hơn (4), tăng tốc độ khử cực pha 4 (5), tốc độ phóng xung nhanh hơn. C, tự động tính bất thường do sự thay đổi hình dạng điện thế hoạt.
VÒNG VÀO LẠI
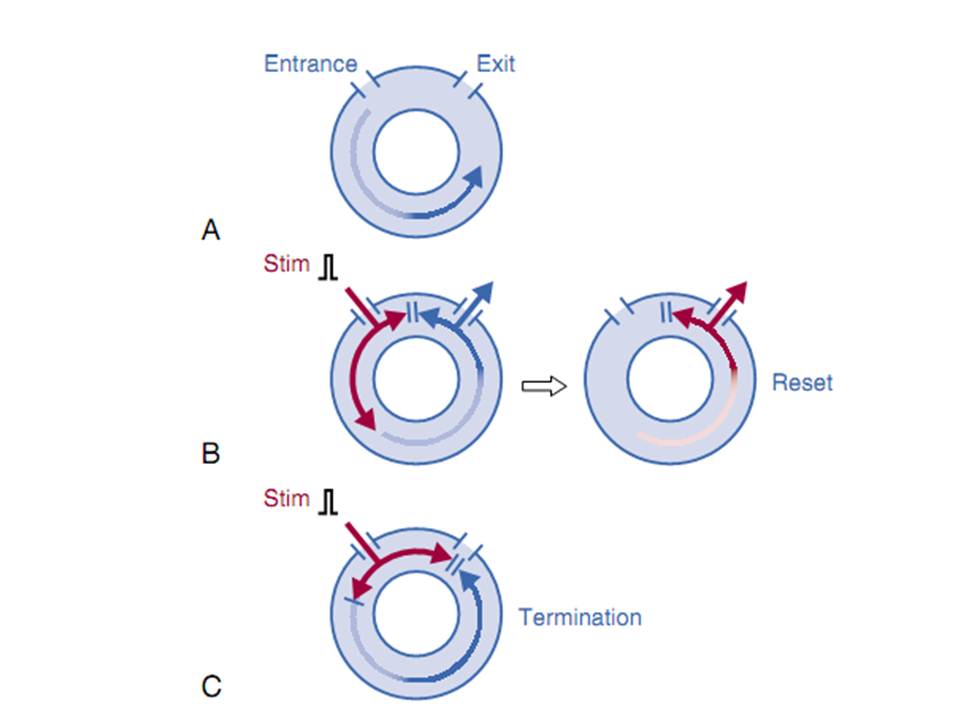
Vòng vào lại là cơ chế loạn nhịp thường nhất là do rối loạn dẫn truyền xung. Cơ chế này đòi hỏi hai con đường khác nhau cho dẫn truyền xung. Các con đường này khác nhau về giải phẫu hay chức năng. Loạn nhịp này xảy ra khi có một xung đến sớm. Kích thích đến sớm này bị blốc trong một con đường và dẫn truyến chậm trên con đường còn lại. Xung di chuyển đủ chậm để cho con đường bị blốc hồi phục và dẫn truyền ngược qua con đường bị blốc gốc đầu tiên. Một nhịp đơn của vòng vào lại được gọi là nhịp echo hay là nhịp dội ngược.Sự duy trì của vòng này tạo ra nhịp nhanh. Độ dài sóng của vòng vào lại bằng vận tốc dẫn truyền nhân với giai đoạn trơ dài nhất của vòng.
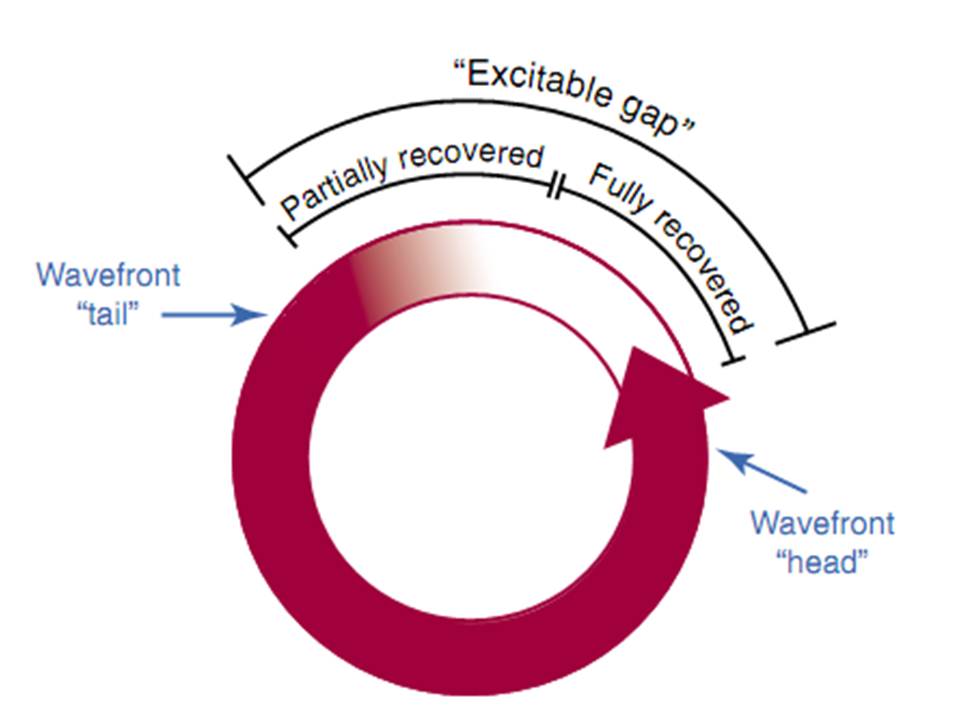
Chất nền giải phẫu của vòng phải đủ lớn để bao cả độ dài sóng. Nếu độ dài chất nền lớn hơn độ dài sóng thì có một khoảng không gian hay thời gian giữa đầu và đuôi của xung kế tiếp được gọi là khoảng ghép kích thích được. Khoảng ghép kích thích được đại diện cho mô không bị trơ và vì vậy có khả năng được hoạt hoá trong nhịp nhanh. Tạo ra một kích thích mà xâm nhập khoảng ghép kích thích sẻ làm thay đổi nhịp nhanh hay chấm dứt nhịp nhanh. Thay đổi nhịp nhanh là do tương tác giữa sóng đến sớm và nhịp nhanh đưa đến xung kế tiếp của nhịp nhanh sẻ đến sớm hoặc trì hoãn. Trong vòng xung đến sớm sẻ vào khoảng ghép kích thích và có hai phần, một phần đối đầu với xung dẫn truyền ngược của nhịp nhanh trước và phần kia sẻ dẫn truyền qua khoảng ghép kích thích để tạo ra một phức hợp đến sớm.Thay đổi vòng vào lại với sự hợp nhất ngụ ý rằng cơ chế vòng vào lại với đường vào và ra tách biệt nhau. Sự hợp nhất có thể biểu hiện trên điện tâm đồ bề mặt hay điện tim khu trú trong buổng tim.
Sự thay đổi liên tục của nhịp nhanh được gọi là entrainment, một nghiệp pháp được sử dụng để chứng minh tồn tại vòng vào lại.

Các kiểu của vòng vào lại. Vùng đậm là mô trơ hoàn toàn, vùng lốm đốm là mô trơ một phần. Vòng vào lại giải phẫu được quyết định do cấu trúc hay mô sẹo của tim và một phần của vòng đã hồi phục kích thích hoàn toàn để có thể bị kích thích trong khi chờ xung kế tiếp đến. Với vòng vào lại chức năng tất cả các phần của vòng phải hồi phục.
CƠ CHẾ LOẠN NHỊP LÂM SÀNG THƯỜNG GẶP
| Vào lại | Hoạt động khởi kích | Tự động tính |
| Nhịp nhanh nhĩ (khởi phát bằng ngoại tâm thu nhĩ) AVNRT AVRT Cuồng nhĩ Nhanh thất do sẹo Nhịp nhanh thất vô căn |
Nhịp nhanh nhĩ (kịch phát bị blốc , ví dụ ngộ độc digoxin gây DAD RVOT VT Hội chứng QT dài (EADs) |
Nhịp nhanh nhĩ (tăng từ từ và kéo dài ở bệnh nhân trẻ) Nhanh thất sau tái tưới máu Nhanh thất do thiếu máu Nhanh thất phân nhánh |
AVNRT: nhịp nhanh vào lại nút nhĩ thất, AVRT: nhịp nhanh do vào lại nhĩ thất, RVOT VT: nhịp nhanh thất từ đường ra thất phải.
Tài liệu tham khảo
- Issa, Miller, Zipes. Clinical arrhythmology electrophysiology. 2009.
- Peter J. Zimetbaum, Mark E. Josephson. Practical clinical electrophysiology. 2009
Tác giả bài viết: TS BS Tôn Thất Minh (BV Tâm Đức)